
Living systems, space, and coherence
Living systems do not operate in a single space in the sense of a bounded domain, nor do they truly inhabit many distinct ones; rather, they instance a continuous relational field whose articulation into behavioural, transcriptional, morphological, physiological, and symbolic sub-spaces constitutes its harmonic structure, not a reduction of it. These sub-spaces are not fragments of a whole that has been broken apart, but resonant modes of a single superspace, each expressing constraints, affordances, and continuities that cannot be collapsed into any other without loss.

The problem of many spaces
Contemporary biology, cognitive science, and systems theory increasingly describe living systems as operating across multiple, partially incommensurable spaces: behavioural space, transcriptional space, morphospace, physiological state space, ecological niche space, and others. Michael Levin’s work on bioelectricity and collective cellular intelligence has been especially influential in making this plurality explicit, arguing that biological systems exhibit goal-directed competence across diverse representational domains rather than being exhaustively described by local molecular causation.
These spaces are not metaphors. They are operational domains in which systems act, stabilise, and correct error. Cells navigate transcriptional landscapes, tissues explore morphospaces, organisms traverse physical environments, and nervous systems regulate physiological manifolds. Each domain has its own coordinates, constraints, attractors, and modes of failure. Yet when these domains are treated as independent levels or stacked layers, an explanatory deficit appears: no single space explains how coherence is preserved across the whole.
This document advances a deliberately unorthodox claim. These domains are not parallel worlds or nested containers. They are harmonic articulations of a deeper, unified superspace. That superspace is not physical, semantic, or symbolic in isolation. It is logical, relational, and non-orientable. I refer to it as the logical orbit.
Levin, information, and form
Levin’s central provocation is that biological systems store and act upon information that is not reducible to genes, synapses, or chemical gradients alone. Bioelectric networks encode pattern memories; tissues pursue anatomical setpoints; cellular collectives solve problems in spaces that resemble planning rather than reaction.
Crucially, these informational structures behave like invariants. A salamander limb regenerates to a target morphology despite perturbations. A planarian reconstitutes its body plan after radical resection. These behaviours indicate the presence of attractors that function analogously to Platonic forms: not as transcendent ideals, but as distributed constraints that guide system dynamics across disturbance.
What remains unresolved is the ontological status of these invariants. They are described as goals, memories, or encodings, while the deeper question remains unanswered: how do multiple informational spaces remain mutually coherent without collapsing into a single privileged description? The logical orbit is proposed here as the missing relational condition.
Sub-spaces as projections of a continuum
The core move is simple but demanding. Behavioural space, transcriptional space, morphospace, and physiological space are not stacked levels. They are projections of a continuous relational field into distinct coordinate systems.
Each projection preserves some relations while obscuring others. Behavioural space preserves trajectories and temporal ordering. Transcriptional space preserves regulatory adjacency and stability basins. Morphospace preserves geometrical feasibility under developmental constraints. Physiological space preserves homeostatic trajectories and compensatory loops.
What is conserved across all projections is not substance, signal, or representation, but structured difference.
Logical antisymmetry and spectral coupling
By logical antisymmetry I mean the following: coherence in complex systems is maintained not by identity, equivalence, or closure, but by structured difference that cannot collapse without destroying the system.
At every scale, systems are sustained by paired descriptions that remain coupled while refusing identity. These pairings form a spectral coupling. They do not merge. They do not map cleanly onto one another. They constrain each other across a persistent gap.
The coupling is real because each side limits and shapes the other. The coupling is spectral because it is expressed as phase, lag, resonance, and interference rather than pointwise translation. The relation persists only so long as difference is conserved.
If the pair becomes identical, the field between them dissolves. Control vanishes. Meaning collapses. The system ceases to exist as a system.

The orbit frame
The orbit frame is the minimal stabilised expression of the logical orbit. It consists of two mutually defining poles and the relational field they sustain. Neither pole is primary. Each exists only insofar as it is not the other.
The orbit frame is a convenience. It is a handle. It is not the ontology. It allows precise discussion of collapse, coherence, and persistence without mistaking the representation for the field itself.
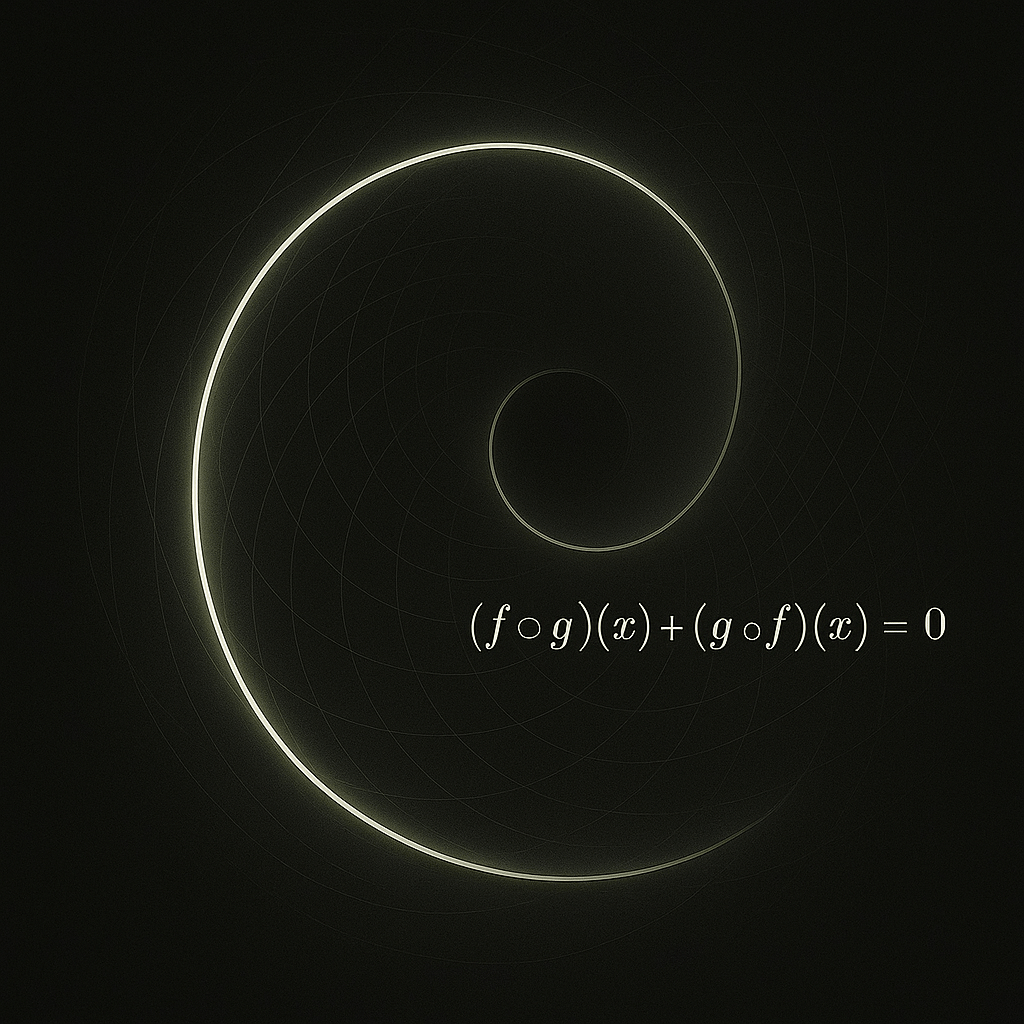
The antisymmetry condition shown here is not algebra to be solved. It marks a boundary: when the coupled descriptions cancel into equivalence, the generative field collapses.
H as a field condition
H names the field condition itself: the irreducible relational tension that cannot be carried by either pole alone. It is not additive. It is not representational. It does not converge.
When written in a self-negating form, the notation does not indicate computation. It signals persistence through recursive self-negation rather than through convergence to identity.
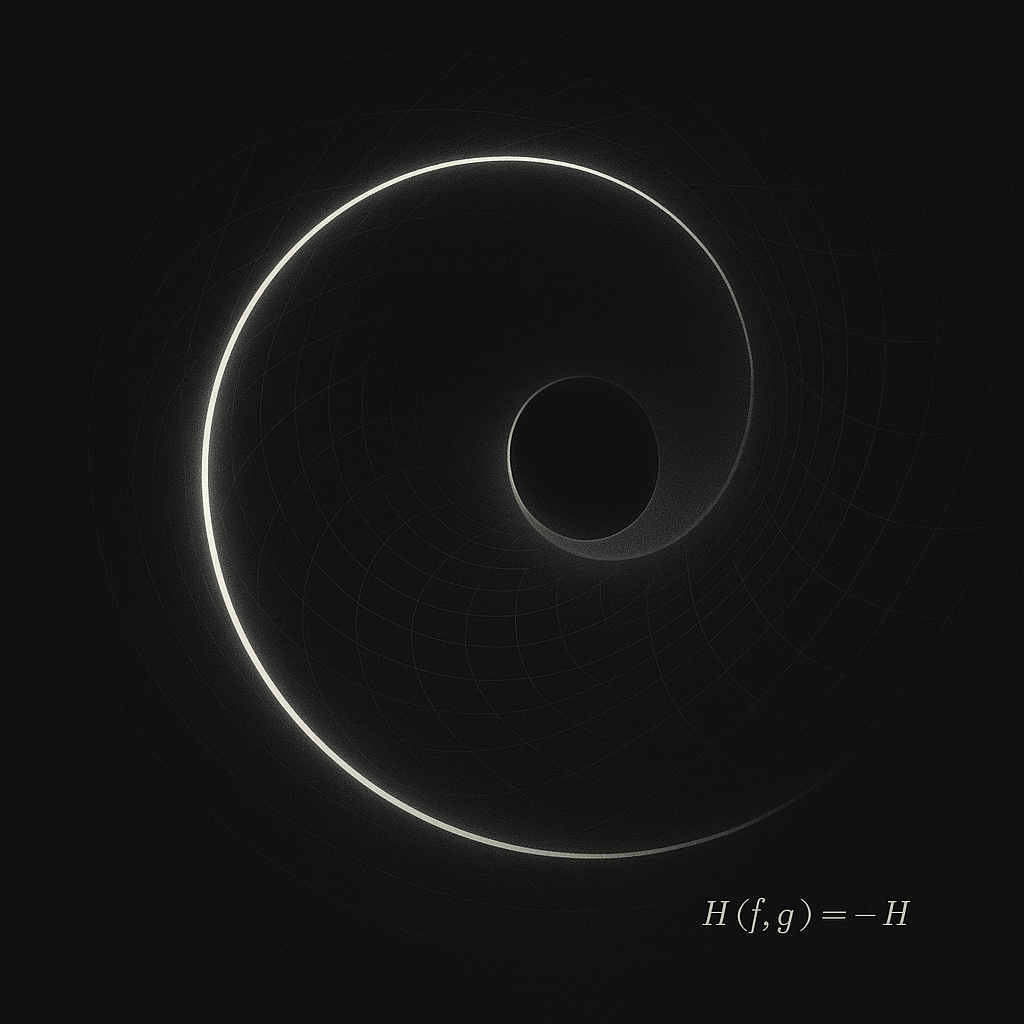
This notation functions as a warning label rather than a derivation. The field persists only because it never resolves.
Experience, language, and priority
These ideas were not derived from language models. They precede my engagement with them by decades. They emerged from sustained reflection on systems that fail, persist, adapt, and deceive themselves. Language models have proven useful as compositional instruments, but they do not originate the framework. They are themselves inhabitants of the logical orbit: systems whose coherence depends on unresolved tension between symbol and world.
Language does not describe the logical orbit from outside. It participates in it. Meaning is not contained in language. Language is contained within the relational field that produces meaning.
Incredulity as signal
Unorthodox frameworks reliably attract dismissal framed as methodological concern. This is not unusual. Paradigms are self-protective systems. They optimise for internal coherence, not external truth. When a framework exposes foundational blind spots, noise often substitutes for engagement.
This is not a claim of persecution. It is a structural observation.
Why this matters
The stakes are not academic. Biological engineering, artificial intelligence, governance, and planetary-scale systems increasingly operate beyond the reach of linear control models. Treating subsystems in isolation accelerates failure. The logical orbit offers a way to think rigorously about coherence without closure, control without domination, and intelligence without centralisation.
If future systems are to remain aligned with the conditions that sustain life rather than erode it, they must be designed with an understanding of antisymmetric relational structure.
Conclusion
The logical orbit is proposed as a unifying superspace underlying the diverse operational spaces identified in contemporary biology and systems science. These spaces are harmonic articulations of a single continuum. Their coherence is conserved by spectral coupling and logical antisymmetry, not by reduction to identity.
This is not a finished theory. It is a scaffold.
If it provokes discomfort, that is expected.
If it provokes careful thought, it has already begun its work.
—
References
Levin, M. (2019). ‘The computational boundary of a “self”: developmental bioelectricity drives multicellularity and scale-free cognition’, Frontiers in Psychology, 10, 2688.
This paper argues that biological systems exhibit cognition-like behaviour across scales, with bioelectric networks enabling goal-directed activity that cannot be reduced to genetic or molecular mechanisms alone. It supports the claim that living systems operate in multiple informational domains simultaneously, without any single domain being ontologically primary. In the context of the logical orbit, this establishes that coherence must be preserved across distinct but coupled spaces, rather than enforced through reduction or hierarchy.
Levin, M. (2020). ‘Life, death, and self: fundamental questions of primitive cognition viewed through the lens of body plasticity and synthetic organisms’, Biochemical and Biophysical Research Communications, 564, pp. 114–133.
This work examines how biological identity and agency persist through radical perturbation, regeneration, and synthetic reconfiguration, challenging fixed notions of organismal selfhood. It is relevant here because it shows that identity is maintained through constraint and regulation rather than static structure. This directly implicates the logical orbit as a relational condition under which persistence is possible without identity collapse.
Levin, M., Keijzer, F. and Lyon, P. (2021). ‘Uncovering cognitive similarities and differences, conservation and innovation’, Philosophical Transactions of the Royal Society B, 376(1820), 20190747.
This paper situates cognition as a conserved but transformable phenomenon across biological systems, emphasising continuity without equivalence. It aligns with the argument that different descriptive spaces can remain coupled without being reducible to one another. Within the logical orbit framework, this supports the notion of spectral coupling: coherence through difference rather than through unification.
Waddington, C.H. (1957). The Strategy of the Genes. London: Allen & Unwin.
Waddington introduces the epigenetic landscape to describe development as movement through a constrained space of possibilities shaped by genetic, cellular, and environmental factors. This provides an early systems-level account of how form and stability emerge without linear causation. Its relevance here lies in anticipating the idea that biological trajectories are guided by higher-order constraints that resemble projections of a deeper relational structure.
Ashby, W.R. (1956). An Introduction to Cybernetics. London: Chapman & Hall.
Ashby formalises the study of regulation, stability, and control in complex systems, introducing principles such as requisite variety and adaptive equilibrium. His work underpins the claim that control depends on maintained difference between regulator and regulated, not on their convergence. In the present argument, this reinforces the necessity of logical antisymmetry as a condition for system persistence.
—
Coda
Underlying everything discussed here is a self-consistent topological space that is neither defined nor formalised in this text. Its existence is assumed, its effects are traced, and its constraints are felt throughout. It is understood rather than specified. The difficulty of explaining it is not a failure of effort, but a consequence of describing something whose existence precedes the descriptions available to it.


One reply on “The Problem of Many Spaces”
LikeLike